INTRODUCTION
Osteoclasts are unique cells that resorb old bones.[1] Osteoclasts play critical roles in many bone-destructive diseases such as osteoporosis, osteomyelitis, rheumatoid arthritis, and periodontitis.[1,2] Therefore, many researchers consider osteoclasts clinically important and have attempted to develop new drugs targeting osteoclasts to treat osteolytic diseases.[3]
Osteoclasts originate from monocyte/macrophage lineage cells.[4] Macrophage colony-stimulating factor (M-CSF) and receptor activator of nuclear factor-κB (NF-κB) ligand (RANKL) are key factors in osteoclastic differentiation.[5] Osteoblasts, bone-forming cells, also regulate osteoclast differentiation via RANKL and osteoprotegerin (OPG) expression, a decoy receptor for RANKL.[5,6] Stimulation of osteoclast precursors with RANKL recruits cytoplasmic tumor necrosis factor receptor-associated factors (TRAFs) and activates downstream signaling pathways. NF-κB, mitogen-activated protein kinases (MAPKs; p38, c-Jun N-terminal kinase, extracellular signal-regulated kinase), and phosphoinositide 3-kinase/Akt serine/threonine kinase (Akt) signaling pathways are known to be activated during osteoclast differentiation.[7-9] Activation of these signaling pathways induces the expression of transcription factors such as c-Fos and nuclear factor of activated T cells (NFATc1), a master transcription factor for osteoclastogenesis.[10-12] c-Fos also induces the expression of NFATc1, contributing to the induction of osteoclast-specific marker genes such as tartrate-resistant acid phosphatase (TRAP).[13,14]
Although several medications targeting osteoclasts have been developed, concerns regarding their side effects have attracted the attention of many researchers.[15-17] The genus Iris belongs to the Iridaceae family and comprises 300 species of perennial flowering plants. Since Iris species display numerous therapeutic effects in folk medicine, they have been used in the treatment of various diseases such as colds, cancer, inflammation, and infections.[18] Among Iris species, Iris Koreana NAKAI (IKN) has a restricted geographical distribution and is endemic to Korea.[19,20] Because IKN has been designated as “threatened to extinction: the first grade (I) for preservation”,[21] its pharmaceutical effects have not been investigated. In this study, we demonstrated the effects of an ethanol extract of IKN roots on osteoclast differentiation in vitro and in vivo. Our study will aid in understanding the therapeutic value of IKN in bone metabolism.
METHODS
1. Reagents
Ethanol extracts of IKN and Iris germanica L. were provided by the National Institute of Horticultural and Herbal Science (Jeollabuk-do, Korea). Briefly, the roots or aerial portions of plants were extracted with 99.99% ethyl alcohol at 85°C using an accelerated solvent extractor. The extract was filtered and concentrated using a rotary evaporator (JP-SD1000; Eyela, Tokyo, Rikikatai, Japan). The final extract was dissolved in dimethyl sulfoxide (DMSO; Sigma-Aldrich, St. Louis, MO, USA) and diluted in phosphate-buffered saline (PBS). Antibodies against p38, phosphorylated p38, β-actin, and c-Fos were purchased from Cell Signaling Technology (Danvers, MA, USA). An antibody against NFATc1 was obtained from Santa Cruz Biotechnology (Dallas, TX, USA). Ultrapure lipopolysaccharide (LPS) from E. coli 0111:B4 was purchased from InvivoGen (San Diego, CA, USA). All other reagents were purchased from Sigma-Aldrich.
2. Co-culture system
All animal experiments were performed in accordance with the National Institute of Health Guide for the Care and Use of Laboratory Animals and the Association for the Assessment and Accreditation of Laboratory Animal Care of Sookmyung Women’s University. Primary calvarial osteoblasts were extracted from the calvariae of neonatal ICR mice (Samtako Inc., Osan, Korea), as previously described.[22] Bone marrow cells were extracted from the long bones of 4 to 6-week-old male ICR mice. To examine osteoclast differentiation, mouse bone marrow cells (1×105 cells) were co-cultured with calvarial osteoblasts (5×103 cells) and 1,25-dihydroxy-vitamin D3 (1,25[OH]2D3; 10 nM) in the presence or absence of IKN root ethanol extract in 96-well culture plates (Corning, MA, USA). After 6 days of culture, the cells were fixed and stained with TRAP staining solution (0.01% naphthol AS-MX phosphate [Sigma-Aldrich] and 0.06% Fast Red Violet LB Salt [Sigma-Aldrich] in 50 mM sodium tartrate dehydrate and 45 mM sodium acetate, at pH 5.0). TRAP-positive (TRAP+) multinucleated cells (>3 nuclei/ cell) were considered mature osteoclasts.
3. Bone marrow macrophage (BMM) culture system
Bone marrow cells were extracted from the long bones of 8 to 10-week-old ICR mice (Samtako Inc.). Bone marrow cells were cultured for 3 days in the presence of M-CSF (30 ng/mL; PeproTech Inc., Rocky Hill, NJ, USA) to generate BMMs. To assess osteoclast differentiation, BMMs were treated with an ethanolic extract of IKN, M-CSF (30 ng/mL), or RANKL (100 ng/mL; PeproTech Inc.). After 4 days, the cells were fixed and stained with TRAP.
4. Cell cytotoxicity assay
Cell viability was determined using the MTT (3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide) assay. BMMs (1×104 cells/well) were seeded into a 96-well plate and incubated with M-CSF (30 ng/mL; R&D Systems, Minneapolis, MN, USA) and the ethanol extract of IKN in α-minimal essential medium for 48 hr. MTT solution was then added, and the incubation was allowed to proceed in the dark. After 5 hr, solubilization buffer (10% sodium dodecyl sulfate [SDS] in 0.01 M HCl) was added and cells were cultured overnight. The viability of BMMs was determined by measuring their optical density at 570 nm.
5. RNA extraction and polymerase chain reaction (PCR) assay
Total RNA was purified using Easy Blue (iNtRON Biotechnology Inc., Seongnam, Korea). cDNA was synthesized from 5 μg of RNA using a RevertAid μFirst-Strand cDNA Synthesis Kit (iNtRON Biotechnology Inc.) and amplified using quantitative PCR (qPCR). Specific primers for genes were as follows: RANKL: 5′-CCA AGA TCT CTA ACA TGA CG-3′ (forward), 5′-CAC CAT CAG CTG AAG ATA GT-3′ (reverse); OPG: 5′-ACG GAC AGC TGG CAC ACC AG-3′ (forward), 5′-CTC ACA CAC TCG GTT GTG GG-3′ (reverse); and GAPDH: 5′-AAC GGA TTT GGT CGT ATT GGG-3′ (forward), 5′-CAG GGG TGC TAA GCA GTT GG-3′ (reverse). Real-time PCR analysis was performed using SYBR® Green PCR Master Mix (Applied Biosystems, Foster City, CA, USA) according to the manufacturer’s instructions. Thermocycling was performed using a 7500 Real-time PCR System (Applied Biosystems) with the following cycling conditions: initial hold, 95°C for 10 min; followed by 40 cycles of denaturation at 95°C for 15 sec, annealing at 58°C, and extension at 60°C for 1 min. The index mRNA level was assessed using a threshold cycle value and normalized to glyceraldehyde 3-phosphate dehydrogenase (GAPDH).
6. Western blot analysis
Total cell lysates were separated using SDS-polyacrylamide gel electrophoresis and transferred onto Immobilon-P membranes (Millipore, Bedford, MA, USA). The membranes were blocked with BSA in PBS-Tween (PBS-T); immunostained with anti-phospho p38 (1:1,000), anti-p38 (1:1,000), anti-NFATc1 (1:200), anti-c-Fos (1:1,000), and anti-β-actin (1:4,000); and incubated with a horseradish peroxidase-conjugated secondary antibody (1:5,000). Membranes were developed using an advanced chemiluminescence detection kit (Amersham Biosciences, Buckinghamshire, UK).
7. Actin ring formation assay
Cells were fixed in 10% formalin for 5 min. The cells were permeabilized with ethanol/acetone (1:1) for 1 min at room temperature and dried. Actin rings were stained with rhodamine-conjugated phalloidin (Molecular Probes, Eugene, OR, USA) overnight in PBS at 4°C in the dark. After washing the cells twice with PBS, images were captured using a fluorescence microscope.
8. Bone resorption assay
BMMs were differentiated on dentin slices in the presence of M-CSF (30 ng/mL) and RANKL (100 ng/mL) for 4 days and then treated with IKN ethanol extract for 2 days. The dentin slices were stained with toluidine blue (1 g/mL; J. T. Baker, Phillipsburg, NJ, USA). The number of resorption pits on the dentin slices was counted.
9. Osteoclast formation in vivo
ICR mice (12-weeks-old) were subcutaneously injected with vehicle (PBS) or LPS (0.5 mg) over the calvarial bone on day 0. Mice were also administered daily intraperitoneal (i.p.) injections of an ethanol extract of IKN roots (50 mg/kg) (dissolved in DMSO and corn oil) or vehicle beginning on day −1. On day 6, the mice were euthanized by CO2 asphyxiation, followed by cervical dislocation. Whole calvaria was extracted, fixed in 4% paraformaldehyde for 24 hr, and stained with TRAP. Image analysis was performed using ImageJ software (version 1.32; National Institutes of Health, Bethesda, MD, USA) according to the manufacturer’s protocol.
RESULTS
1. IKN root suppresses osteoclast formation and bone resorption
We first investigated the effect of an ethanol extract of the root part of IKN on osteoclast differentiation using a mouse co-culture system. Mouse bone marrow cells and osteoblasts were cocultured with 1,25(OH)2D3 to induce mature TRAP+ multinucleated osteoclasts (MNCs). We found that the ethanol extract of IKN roots dramatically suppressed osteoclast differentiation (Fig. 1A).
As osteoblasts are known to regulate osteoclast differentiation via the expression of RANKL and OPG,[5,6] we further determined the expression levels of RANKL and OPG in the presence or absence of IKN root ethanol extract. qPCR analysis showed that the addition of 1,25(OH)2D3 increased the mRNA level of RANKL, which was not altered by treatment with IKN ethanol extract (Fig. 1B). Similarly, the OPG mRNA level was decreased by the addition of 1,25(OH)2D3, and the presence of the ethanol extract failed to interfere with this (Fig. 1C). Altogether, we concluded that the anti-osteoclastogenic effect of the ethanol extract of IKN roots may not be mediated by osteoblasts.
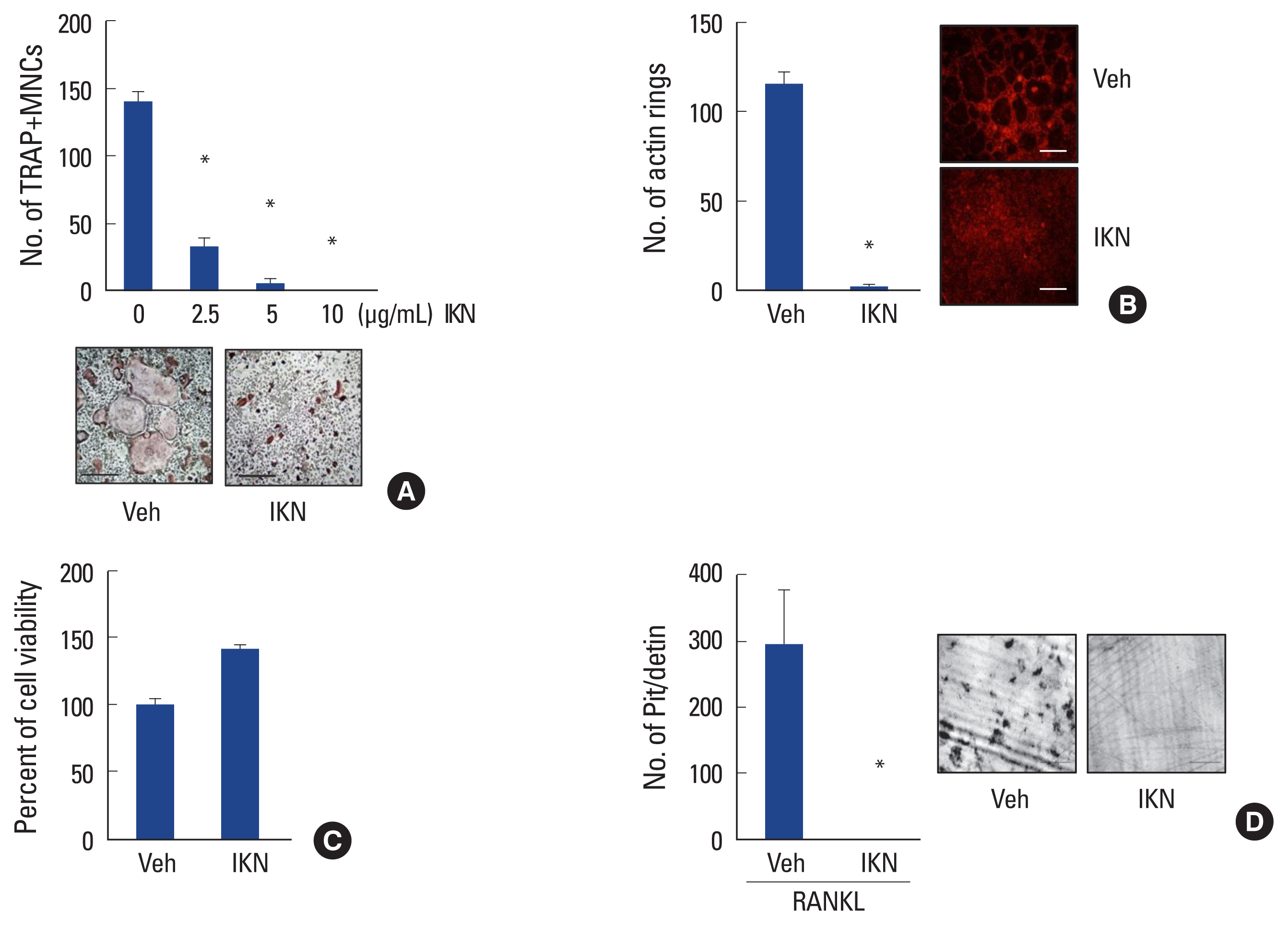
Next, we examined the direct effect of IKN root ethanol extract on mouse bone marrow cells. When mouse bone marrow cells were cultured with M-CSF for 3 days, they differentiated into BMM, which are precursors of osteoclastic cells. Further culture of BMM with RANKL in the presence of M-CSF generated mature TRAP+ MNCs (Fig. 2A).[4,5] We found that treatment with IKN ethanol extract failed to induce the differentiation of BMM cells into TRAP+ MNCs, even in the presence of RANKL. The anti-osteoclastogenic effect of the IKN ethanol extract was observed in a dose-dependent manner with a maximum concentration of 10 μg/mL (Fig. 2A).
Mature osteoclasts primarily function as bone-resorbing cells. To efficiently resorb bone, osteoclasts form an F-actin ring to generate a sealing zone.[23] Thus, we examined the effect of IKN root ethanol extract on F-actin ring formation during osteoclastogenesis. We observed F-actin ring structures in cultures of BMMs with RANKL; however, treatment with IKN ethanol extract inhibited the formation of F-actin rings (Fig. 2B).
To address whether the anti-osteoclastogenic effect of the IKN root ethanol extract was due to its cellular toxicity, we performed a cell viability assay using the maximum concentration of the IKN root ethanol extract. Compared to the vehicle, treatment with IKN ethanol extract did not impair cell viability (Fig. 2C). These data suggested that the inhibitory effect of the ethanol extract of IKN roots was not attributed to cellular toxicity.
Because the ethanol extract of IKN root exhibited a suppressive effect on the formation of osteoclasts and F-actin rings in vitro in plastic plates, we further investigated whether the anti-osteoclastogenic effect of the ethanol extract of IKN root was associated with the bone-resorbing activity of osteoclasts. BMMs cultured on dentin slices in the presence of RANKL-generated resorption pits. However, treatment with IKN ethanol extract efficiently reduced the number of resorption pits on dentin slices (Fig. 2D). Taken together, these results suggested that the ethanol extract of IKN root suppresses osteoclast differentiation, leading to the inhibition of bone resorption.
2. Root part of IKN suppresses NFATc1 and c-Fos protein induction possibly via the p38 MAPK pathway
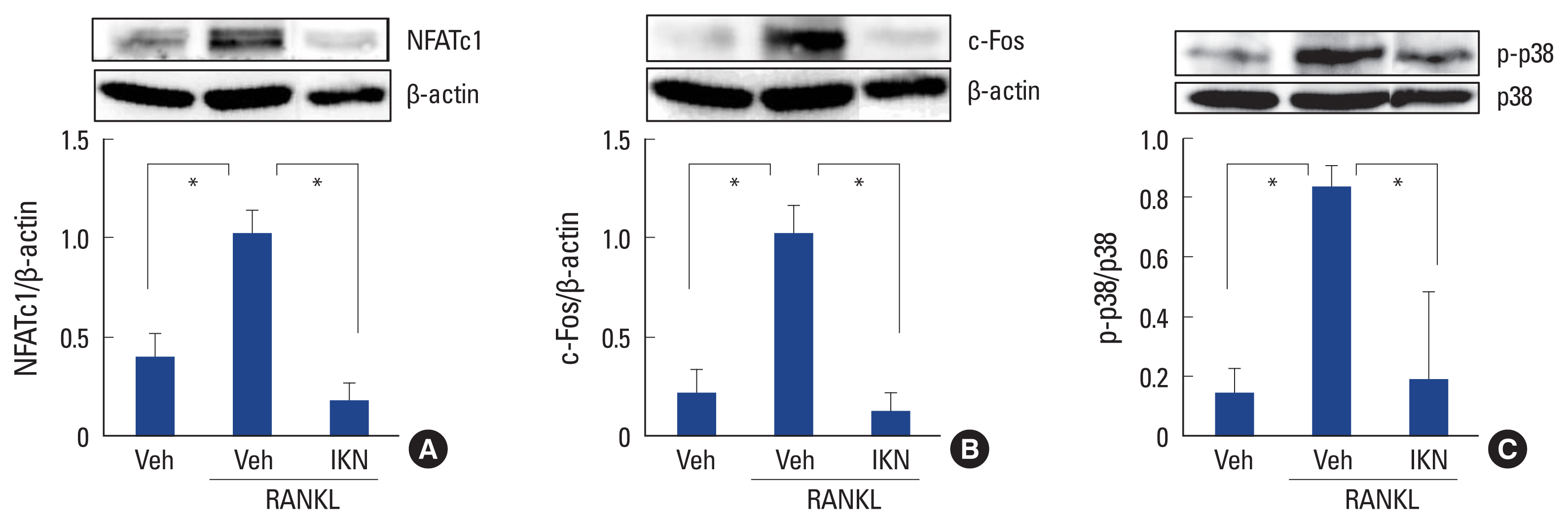
We verified that the ethanol extract of IKN roots had inhibitory effects on RANKL-induced osteoclast differentiation in BMM cell cultures. To clarify the molecular mechanisms involved, we investigated whether the ethanol extract of IKN roots affects signaling pathways involved in osteoclast differentiation. Because NFATc1 is a master transcription factor for RANKL-induced osteoclast formation, [11,12] we initially measured the expression levels of NFATc1. Culturing BMMs with RANKL significantly induced NFATc1 protein expression, which was completely abolished by treatment with IKN ethanol extract (Fig. 3A). c-Fos is widely known to positively regulate NFATc1 and osteoclast differentiation.[10] When we further investigated the protein expression levels of c-Fos, the IKN ethanol extract suppressed RANKL-induced expression of c-Fos (Fig. 3B). These findings suggested that the ethanol extract of IKN roots modulates osteoclast differentiation by suppressing the induction of c-Fos and NFATc1.
Next, we explored the upstream signaling pathways of c-Fos and NFATc1 in osteoclast formation. We found that treatment with the ethanol extract of IKN roots significantly inhibited the phosphorylation of the p38 MAPK pathway by RANKL in BMM cells (Fig. 3C). Given that the p38 MAPK pathway is activated via phosphorylation, the ethanol extract of IKN root of IKN appears to inhibit the p38 MAPK pathway, which leads to impaired osteoclast differentiation.
3. Anti-osteoclastogenic effect is unique to the root part of IKN
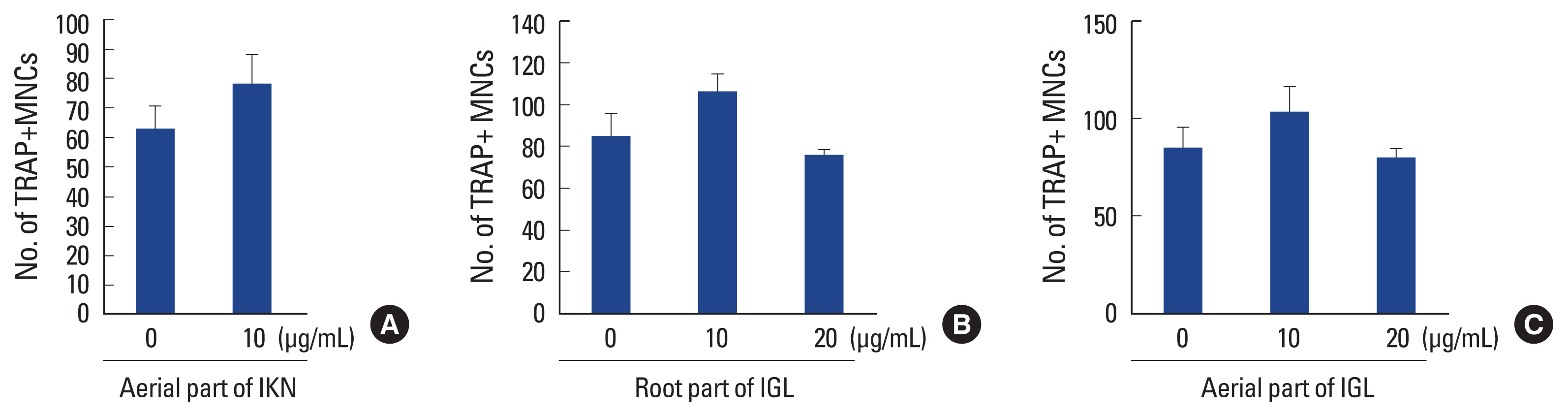
In addition to the underground components, the aerial parts of Iris species are a rich source of bioactive ingredients.[18] Therefore, we investigated the effect of an ethanol extract of the aerial part of IKN on osteoclastogenesis. In contrast to the root part of IKN, the ethanol extract of the aerial part of IKN failed to interfere with RANKL-induced osteoclast formation in BMM cells (Fig. 4A).
Iris germanica L. is one of the most widely investigated Iris species.[24] To verify that the anti-osteoclastogenic effect was a specific property of IKN, we examined the effect of an ethanol extract of Iris germanica L. on osteoclast formation. Treatment with the ethanol extract of the aerial or root parts of Iris germanica L. did not suppress RANKL-induced osteoclast differentiation in BMM cultures (Fig. 4B, C). These results suggested that the ethanolic extract of IKN roots exerts a unique effect on osteoclast differentiation.
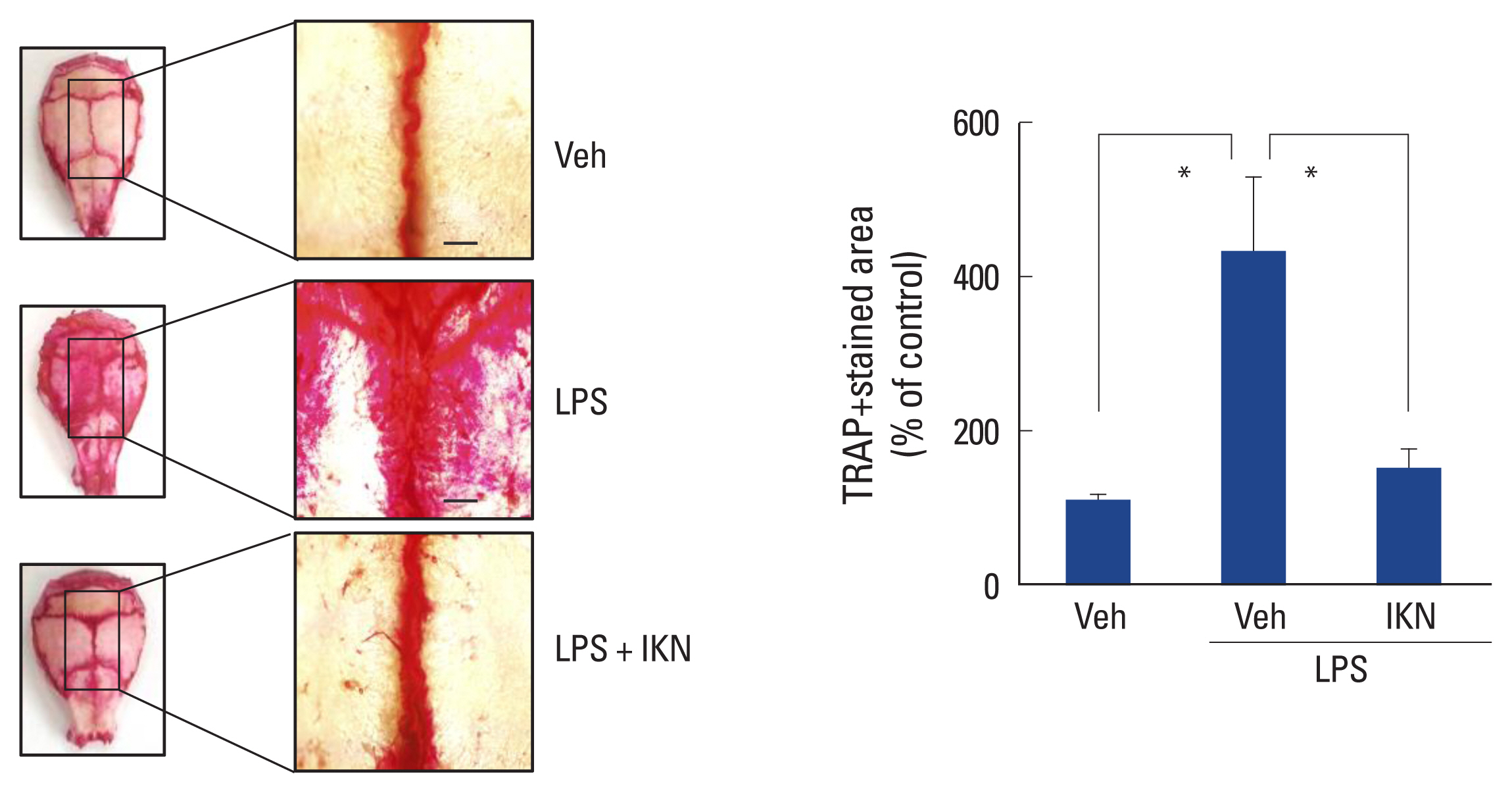
4. Root part of IKN inhibits LPS-induced osteoclast formation in vivo
We confirmed that the ethanolic extract of IKN roots inhibited osteoclast formation in vitro. To examine this in vivo, we used a mouse calvarial bone destruction model.[25] Lipopolysaccharide (LPS) was injected into the subcalvarial area of mice to induce osteoclast formation in vivo. The pretreatment effect of the ethanol extract of IKN root was determined by TRAP staining of whole mouse calvariae. The injection of LPS significantly increased the TRAP-stained osteoclastic area compared to the vehicle (Fig. 5). However, i.p. administration of the ethanol extract of IKN roots prior to LPS dramatically decreased the TRAP-stained osteoclastic area in vivo, which coincides well with the effects observed in vitro. Altogether, these results suggested that the ethanol extract of IKN roots effectively prevented LPS-induced osteoclast formation in vivo.
DISCUSSION
Herbal plants are widely used for the treatment of metabolic bone diseases.[15-17] Because little is known regarding the effects of IKN on bone metabolism, we investigated the effects of IKN on osteoclast differentiation. Osteoblasts induce osteoclast differentiation by upregulating RANKL expression and downregulating OPG expression.[5,6] However, the ethanol extract of IKN root did not alter the expression levels of RANKL and OPG in osteoblasts, suggesting that IKN has little effect on osteoblasts. In a mouse BMM culture system, the ethanol extract of IKN root directly suppressed RANKL-induced osteoclast formation and bone resorptive activity. In contrast, the ethanol extract of the aerial parts of IKN had only a minor effect on RANKL-induced osteoclast differentiation. The ethanol extract of IKN roots was further demonstrated to downregulate the p38 MAPK and c-Fos-NFATc1 signaling pathways. The p38 MAPK signaling pathway plays a positive role in osteoclast formation and bone resorption.[26-29] During osteoclast differentiation, p38 MAPK is recruited to the tumor necrosis factor-receptor-associated factor 6-transforming growth factor-β-activated kinase 1 complex [29] and phosphorylated. Activated p38 MAPK directly phosphorylates and stimulates NFATc1, a master transcription factor essential for osteoclastogenesis, and induces several osteoclast-specific marker genes, such as TRAP and cathepsin K.[30,31] The use of pharmacological inhibitors or siRNAs specific for p38 MAPK is well known to suppresses RANKL-induced osteoclast formation.[32,33] Thus, we assume that downregulation of the p38 MAPK pathway by the ethanol extract of IKN roots may partly contribute to impaired osteoclast formation. However, the precise molecular mechanisms underlying the suppressive effect of the root part of IKN on osteoclastogenesis requires further investigation.
Iris species have numerous pharmacological effects and several dietary supplements; pharmaceutical medicines using Iris species are available commercially on the market. For example, the dried extract of Iris lactea Pall. is widely used as an anti-inflammatory agent to reduce the side effects of chemotherapy and radiation.[34] The rhizomes of I. Versicolor is used to treat mastopathy and menstrual disorders.[35] In addition, various phytochemical compounds of Iris species have been identified and shown to have anticancer, antioxidant, antiplasmodial, immunomodulatory, and anti-inflammatory activities.[18] Previously, an extract of Iris pseudacorus was shown to diminish RANKL-induced osteoclast formation using RAW 264.7 murine macrophages.[36] However, the pharmaceutical effects and bioactive compounds in IKN have not been explored. In this study, we demonstrated a novel effect of IKN root extract on osteoclast differentiation in vitro and in vivo. The anti-osteoclastogenic effect of the ethanol extract of IKN root was not shared by the ethanol extract of Iris germanica L., suggesting a unique effect of IKN. Our study had certain limitations regarding identification of the specific anti-osteoclastogenic component of the ethanol extract of IKN roots. Thus, it would be worthwhile to isolate and verify the bioactive components of the IKN root ethanol extract for the prevention of osteolytic diseases.
IKN is thought to have originated from allopolyploidization between I. minutoaurea and I. odaesanensis.[37-39] Owing to its rarity and restricted distribution, its therapeutic value has not yet been fully investigated. To overcome this rarity, an efficient in vitro IKN propagation method was explored.[20] Mass propagation through in vitro multiplication protocol enhances the application scope of IKN.
In conclusion, we demonstrated that an ethanol extract of IKN roots inhibited RANKL-induced osteoclast differentiation. The suppressive activity of the root component of IKN partly involves p38 MAPK-mediated NFATc1 expression. As many effective drugs from plants are available, the root part of IKN could represent a promising candidate for new therapeutics targeting osteoclast-related bone diseases.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print