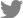INTRODUCTION
The use of allogenic human grafts harvested from femoral heads during hip prosthetic surgery or from a cadaveric donor are common,[1-3] it is a valuable option because of its biophysical properties, micro- and macro-architecture. However, the procedure used to make the grafts safe impairs the osteoinductive capacity of the bone structure and, as already reported, integration with surrounding tissue was limited and the consolidation process slow compared to autograft.[4]
The integration of inorganic bone matrix is related to the local environment and particularly to there being sufficient numbers of osteoprogenitor cells.[5] The demonstrated ability of stem cells in adult bone marrow (BM), termed mesenchymal stem cells (MSCs), to differentiate into osteoblasts, chondrocytes, muscle cells, and stromal adipocytes offers an approach for improving the integration of bone grafts to deal with extensive bone loss.[6-8]
For in vitro colonization of allogeneic bone matrix (ABM) by autologous MSCs prior to grafting - a procedure we have termed ‘graft personalization’ - we studied the in vitro compatibility of a secured inorganic human bone matrix with MSCs from normal adult BM. Different treatments of the scaffold were tested, including the use of human serum (HS) and we studied the adhesion to the bone matrix.
METHODS
1. Human normal BM cell source and isolation
Human BM cells were obtained from 17 femoral heads collected during hip arthroplasty. This was carried out according to a protocol approved by the Regional Ethical Committee. All donors (mean age, 63 yearrs) had normal hematopoietic function. Normal BM cells were collected in a syringe containing heparin and transferred to ethylenediaminetetraacetic acid (EDTA)-tubes (5 mM). From each sample, we mechanically dissociated the low-density pre-formed coherent tissue aggregates, which contained about 60% of the whole BM mesenchymal progenitor cell content.[9] The non-adherent BM mononuclear cells were harvested in 2 gentle rinses and retained for sustaining-hematopoiesis assays. Adherent cells were fed by a weekly change of medium (α-MEM 10% FCS, 100 U penicillin, 100 μg/mL streptomycin, 2 mM L-Glutamin). Mesenchymal cells (MCs) were collected by trypsinization. All the Lin-cells, and some MC, were cryopreserved at −80°C for subsequent use.[10] The number of colony-forming unit-fibroblast-like cells (CFU-F) [11,12] in the cell population was assessed by plating a duplicate fraction at low densities (60 and 240 cells/cm2) for 10 days.
2. ABM
Allogeneic bone samples were collected from femoral heads, taken from patients undergoing total hip arthroplasty. They were first secured at a tissue bank (Osteobanque, Clermont-Ferrand, France) and then treated with the Osteopure process (Ost developpement, Clermont-Ferrand, France). The steps were: removal of cortical bone, cleaning under pressure, physico-chemical treatment to decrease the lipid content with acetone and ethanol, 2 urea washes and γ-sterilization.
3. Assessment of in vitro colonization
To enhance MCs adherence, ABM were incubated with 20 μg/mL gelatin (N=11) or 100 μg/mL collagen IV (N=5) or 20 μg/mL fibronectin (N=5) or HS (N=7) for 2 hr. Because of the high variability of trabecular bone-derived matrix in overall surface area, 3 ABM fragments (about 2 mm3 each) per hole were plated in triplicate in 24-hole plates to give a representative series of nine fragments per condition at each time point. Each bone fragment was sowed with the same number of expanded MCs (1.5×104 cells) in a standard medium, which was changed twice weekly. To evaluate the kinetics of MC proliferation in each condition, 6 arms with 3 wells, each containing 3 bone fragments, were seeded on the same day (D0). After 1, 2, 3, 4, 6 and 8 weeks, 3 holes (one arm) were sacrificed to collect cells. Cells were harvested after detachment by trypsin/EDTA 0.25% and counted. For each arm, we evaluated in parallel the kinetics of cell proliferation on equivalently pre-coated plastic holes (gelatin, collagen IV, fibronectin or AB HS) in the absence of ABM as a control in order to estimate the effect of the bone matrix only.
In additional experiments (N=6), we evaluated the effect of initial MC adherence on ABM colonization. We seeded MC in exactly the same conditions but sacrificed 3 wells 18 hr later and counted adherent cells. In 3 experiments, 3 other holes were maintained in parallel, for 3 weeks then also sacrificed and MC counted to assess the effect of initial adherence on MC proliferation.
4. Flow cytometry
We analyzed the size, the granularity of colonizing CD45− CD14− cells and the expression of CD73 antigen to evaluate possible changes in cell characteristics.[13] In all cases, the MCs were first incubated with PBS/5% HS for 15 min, then incubated for 30 min on ice with monoclonal antibodies specific for CD45 (FITC), CD14 (FITC), and CD73-PE (Beckman Coulter, Roissy, France). Cells were washed once in PBS/4% FCS and once again in PBS/4% FCS with 1 μg/mL propidium iodide (PI). Acquisition and analysis of the viable cells (PI-) was performed on an EPICSElite (Coulter Corp., Miami, FL, USA). Gates defining negative cell subsets were set using the PE and FITC - conjugated isotype controls.
5. In vitro hematopoietic supporting function of MCs
In the standard medium we used, MC usually preserve their capacity to sustain hematopoiesis in vitro. We compared long-term culture in the presence of ABM with culture on routine MC monolayer. Non-adherent cells were depleted of cells expressing CD2, CD3, CD14, CD16, CD19, CD24, CD56, CD66b, and glycophorin A by a negative selection process. This was performed in accordance with the manufacturer’s instructions for collecting lineage-negative (Lin−) cells, enriched in primitive hematopoietic cells (Stem SepTM; StemCell Technologies, Vancouver, Canada). We seeded 5×103 BM Lin− CD34+ cells per hole (15×103/arm) on personalized ABM in a long-term culture medium that included IMDM, 12.5% FCS (BioWest, Nuaille, France), 12.5% horse serum (StemCell Technologies), 2 mM L-glutamine, 10−4 β mercaptoethanol and 10−6 M hydrocortisone (Sigma, Saint-Quentin-Fallavier, France). After replacing half the medium once a week over a 6-week period, all the cells from each hole were harvested and plated for the colony-forming cell (CFC) assay, as already described.[9] Monolayers of MC were used as controls and a MS5 cell line as a positive control. We were able to calculate the number of long-term culture initiating cells (LTC-IC) by dividing the total CFC by the average CFC produced from one LTC-IC under the same conditions.[9] In parallel, we assessed the input frequency of LTC-ICs and the number of LTC-ICs maintained for 6 weeks by plating the input and output cells in a limiting dilution assay (LDA) for a further 6 weeks on a MS5 cell line monolayer.
6. In vitro osteoblastic differentiation
In other experiments, we replaced standard medium with medium inductive for osteoblastic differentiation. This contained α-MEM 10% FCS, 10−2M β-glycerol, 5×10−3M ascorbic acid and 10−8M dexamethasone [14,15] and was done at D0, at the end of the exponential phase of proliferation (week 3) and for 3 additional weeks. MCs were then collected after 1 hr incubation with collagenase and assessed for their alkaline phosphatase (ALP) activity on cytospin. MCs were scored by 2 blinded independent observers under microscope in four intensities; 0, negative; 1, light beige; 2, regular moderate brown color; 3, intense brown; 4, intense brown nearly black and covering the nucleus. A total ALP score was calculated by adding the score of each MC category (resulting from the percentage of MC) multiplied by the positivity score.
7. Statistical analysis
Data were normalized by an arbitrarily defined number of input cells. Results were expressed as means±standard deviation. The LDA analysis was performed according to the Poisson statistical model (L-CalcTM; StemCell Technologies). The statistical analysis used the bilateral paired Student’s t-test with a significant P value less than 0.05.
RESULTS
1. Isolation and in vitro expansion of BM MCs before use
The number of MCs produced from an average of 5 mL BM sample is described in Table 1.
2. Proliferation of MCs into bone matrix
After depositing the in vitro-expanded MCs from P2 or P3, the ABMs were observed at each medium exchange. We noted the first visible bone-adherent cells during the second week, but the bone opacity hampered finding MC. Two weeks later, the cells had proliferated enough to be seen under microscope in bone pores, becoming organized in 3-dimensional (3D) structures after the third week in all coating conditions (Fig. 1). In eleven experiments, we followed the kinetics of the MC proliferation on gelatin pre-coated-bone matrix, a coating favoring cell adherence. The curve of MC proliferation showed a proliferative phase reaching a maximum at week 4 and finishing with a phase of decrease (Fig, 2A). The normalized numbers of MC collected at W2, W3, and W4 were significantly higher in the presence of bone matrix (P<0.05) resulting in a 2-fold higher amplification (×13.3 vs. 6.6). Then, the 2 curves tended to meet, possibly due to increased senescence in our culture conditions and probably related to the higher cell metabolism indicated by a decreased medium pH. The kinetics of proliferation of MC on bone matrix pre-coated with components with potential for clinical use, i.e. collagen IV (N=5), fibronectin (N=5), or decomplemented AB HS (N=7), were followed. In each case, we expressed the result as the number of MC in the presence of ABM divided by that in control (same coating process but without ABM). The proliferation kinetics were roughly similar to those obtained with gelatin coated-ABM (Fig. 2B). AB HS coating seemed to favor their proliferation with a significantly higher number at W2, W3, and W4, as compared to the control.
3. Role of initial MC adherence in bone matrix colonization
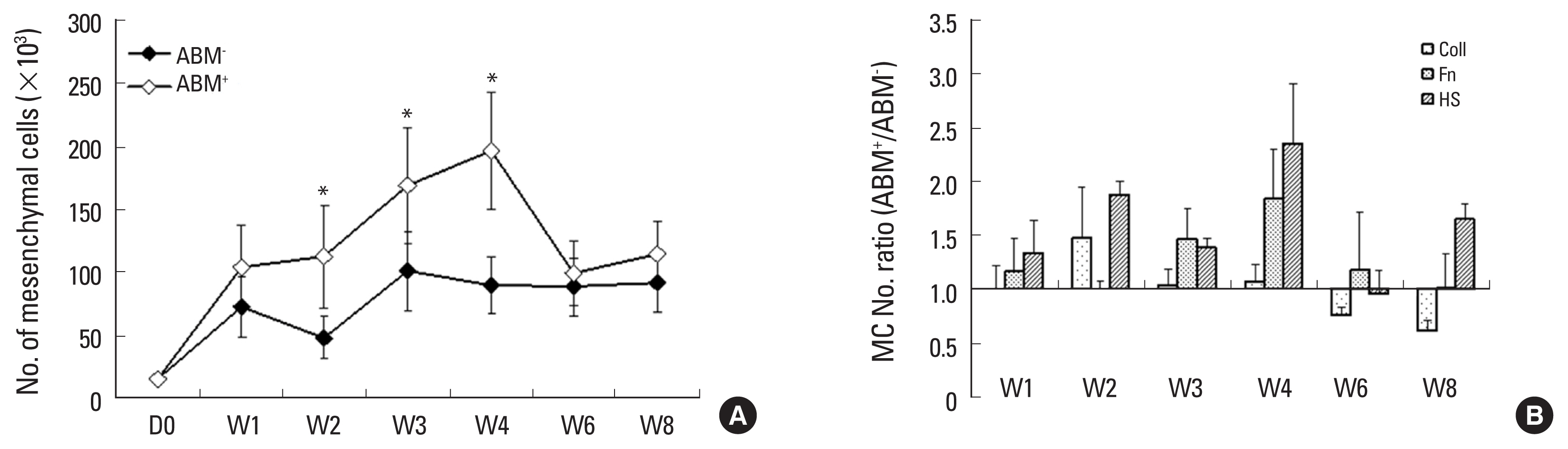
To assess the effect of ABM coating on MC adherence, we compared fibronectin and HS coating with PBS (N=6). After 18 hr of incubation, we observed significantly more adherent cells on HS-coated ABM than on the uncoated ABMs (1.8 fold) or the PBS-incubated (1.7 fold; P<0.05) (Fig. 3A). The difference with regard to fibronectin-coated ABMs was at the limit of significance. Furthermore, when we collected MC from the 3 replicates left to proliferate for 3 weeks (N=3), we found significantly more MC on HS-coated ABM (Fig. 3B) but within a similar range to the initial cell adherence, suggesting that the higher MC production was related to the larger number of adherent input cells.
4. Flow cytometry characterization of ABM-colonizing MC
In the perspective of a clinical application, we checked the characteristics of MC. The CD45− CD14− CD73+ mononuclear subset contains fresh CFU-F.[16] We analyzed ABM-colonizing MC by flow cytometry. We observed significantly smaller cells (P<0.05) in the presence of ABMs, whatever the initial conditioning, compared with cells from the wells without bone matrix until the sixth week (Fig. 4). The ABM+ curves then joined the ABM- curve. Granularity did not appear to be significantly different, except on D42 in the case of gelatin, and we observed fewer CD45− CD14− CD73+ cells on D14 and D21 (fibronectin and AB HS). The mean fluorescence was lower in the 2 first weeks in the presence of ABM pre-coated with AB HS. We noted that size, structure and mean of fluorescence were inversely correlated with the duration of culture.
5. Treated bone did not impair osteoblastic differentiation of colonizing MC
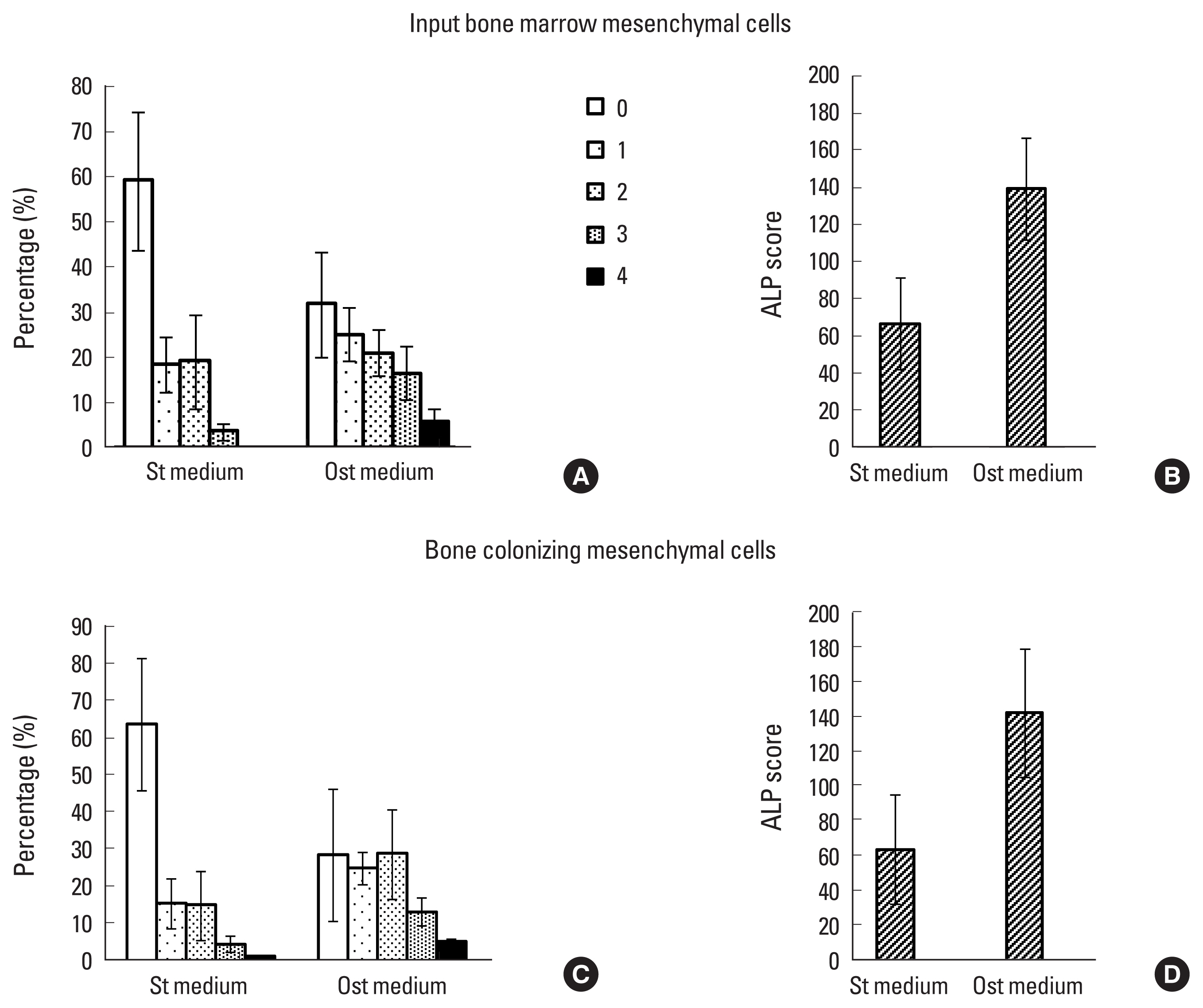
The osteoblastic differentiation of MC was evaluated on D0 (input MC) and after 3 weeks of colonizing ABM. At these time points, cells were incubated in osteoblastic or standard medium for 3 more weeks, then collected and scored for their ALP activity. The osteoblastic medium induced the production of cells with high ALP activity (stage 3 and 4) from the input cell population, mainly at the expense of cells with no activity (stage 0; P<0.03) (Fig. 5A), thus resulting in a 2-fold increase in total ALP in comparison with a standard medium (Fig. 5B). After 3 weeks growth in bone matrix, the MC was cultured under the same conditions. The distribution pattern of cells according to their ALP activity and the increase in total numbers after osteoblast differentiation was very similar to that observed for input cells, demonstrating that osteoblast potential is conserved after colonization with ABMs (Fig. 5C, D).
DISCUSSION
In this study, we evaluated the in vitro ability of normal BM MCs to colonize bone surrogates in a standard medium containing 10% FCS. Under every experimental condition, MC adhered to ABM surface and proliferated with higher amplitude than on the bottom of the control wells and did so by forming 3D structures similar to those described for other scaffolds.[17] This could be due on the one hand to the improved initial adhesion of mesenchymal progenitors since the most effective HS coating induced an increase in cell production, and on the other hand, to the surface area of the bone matrix, which by reducing cell density, could favor cell proliferation.[18,19] The effect of HS may be explained by the adsorption of undetermined molecules (apart from fibronectin),[20] facilitating MC adhesion [21] and was probably related to adhesin-dependent signaling events involved in cell survival and growth.[22] Under all conditions, including the control cultures, we observed similar kinetics of CFU-F production, with a trend to decrease with culture time. However, the presence of ABM did not affect CFU-F numbers and, in this aspect, the cell-to-cell contacts in 3D and cell multilayer did not influence the frequency of progenitors. We noted that bone matrix tended to preserve the initial proliferative potential of CFU-F (colonies >25 cells) for 1 to 2 weeks, that could explain the extra numbers of MC from seeded ABM during the first 4 weeks, particularly with AB HS-coated ABMs. The personalization of grafts is a new concept which aims to obtain a graft adapted to each patient. The final step will be to grow the patient’s cells in the future transplant that will be implanted, our study allows a preliminary step to this project.
The treated bone did not modify the ability of MC to differentiate towards the osteoblastic Lin since the number of osteoblasts-like-cells was similar before and after colonizing ABM. This observation, associated with the maintenance of a significant proportion of CFU-F, suggests that personalized ABM contained progenitors able to proliferate and differentiate in vivo under the influence of local stimulating factors at the fracture site.[23,24] Primitive MC may differentiate towards different cell types depending upon site-specific physiological conditions.[25] However, the fact that ALP activity was not increased by the presence of ABM confirmed that the inorganic ABM is not osteoinductive.
Throughout this study, we noted that MC was maintained in vitro for over 10 weeks. This, and the prolonged phase of cell expansion on bone matrix, suggests that the MC may be able to continue to colonize the bone matrix in vivo. Several teams have been working on the pre-colonization of grafts by undifferentiated MSCs and the approach has been validated in the rabbit,[26] sheep,[27,28] dog,[29] and in man.[30] But these teams used ceramics or coral and not a human bone matrix. However, biomaterial implants are subject to such limitations as inappropriate mechanical properties and poor integration with surrounding tissue.[31] Our approach may appear to go against the current trend by using an allogeneic secured inorganic bone matrix, but the use of this kind of bone surrogate has some evident advantages: (1) its biomechanical properties are very close to those of viable bone [32] offering immediate structural support that may enhance osteogenesis [33]; (2) its porosity and macrostructure should allow graft vascularization; (3) there is no risk associated with xenogeneic graft, comparable to that arising from bovine material, transmission of non-conventional agents (prions) or graft-host incompatibility; (4) their collection is relatively simple. No previous study has tested the biocompatibility of undifferentiated MC and the natural allogeneic inorganic bone matrix and this work is original in this respect. Here, we have validated the first step in the personalization of allogeneic processed-bone matrix.
ABMs coated with autologous serum and colonized by recipient MCs provide an attractive bone bio-graft. Furthermore, we have confirmed that collecting normal BM MCs and performing in vitro expansion in simple medium are feasible procedures and that the expanded MC may be cryo-preserved at −80°C. This is a cheaper and simpler technique than the preservation in nitrogen that we currently use with hematopoietic cells.[34] However, although we obtained a large number of MC in vitro, we noted, as have others, that the CFU-F frequency was low (<5%) and heterogeneous in proliferative capacity.[7,35,36] Furthermore, we observed an early decrease in the CFU-F proliferative potential through planting out; a fact that urges the use of earlier passages for clinical applications.


 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Full text via PMC
Full text via PMC Download Citation
Download Citation Print
Print