INTRODUCTION
The balance between bone growth factors and bone resorption factors, which plays a central role in bone morphology, is constantly adjusted through bone metabolism.[1] Bone metabolism is affected by a variety of factors, including hormones,[2] reactive oxygen species,[3,4] and the autonomic nervous system.[5] Moreover, these bone-associated factors act in a synergistic or conjugated manner depending on exercise loading and internal body conditions such as secretions to form a more complex bone metabolism mechanism.[6,7,8] In short, we believe that the influences on bones differ depending on the internal body environment, which differs between the growth phase and adulthood, as well as on external mechanical influences. The role of osteocytes as a bone remodeling control mechanism has gained attention recently, and an association between bone remodeling and exercise loading has been suggested.[9,10,11] Mechanical influences on the bone can be broadly classified into mechanical stress and strain and are involved in the expression of sclerostin, an inhibitory factor of bone growth.[12]
Kamel et al.[13] and Price et al.[14] support the hypothesis that stress responses induce stimulation of the bone. They reported that increased application of stress to the bone leads to osteogenesis and that mechanical stress influences bone mass and strength in accordance with Wolff's law and the mechanostat model.[15,16,17,18,19] Researchers have confirmed that exercises like jumping exert a stronger impact on the bones than exercises such as swimming that have minimal impact. In other words, identifying the bone metabolism responses to continuous exercise stress during the growth period may be the key to elucidating the acquisition of peak bone mass (PBM) up to adulthood. However, the dynamics of bone metabolism, by which the type of exercise stress applied in the in vivo environment during the growth stage determines bone mass and strength, have yet to be elucidated. The influence of type of exercise stress, exerted post-growth period, on bone strength and mass in adulthood is also not known.
In this study, therefore, we examined the influence of exercise stress type that is continuously exerted since the growth period on bone metabolism. Another goal was to study the influence of bone metabolism responses that are brought about by the specific characteristics of exercise stress on bone strength and growth, and to identify the ideal form of exercise stress that is likely to be required during the growth period.
METHODS
This research obtained and carried out consent of the Ethics Committee of Aomori University of Health and Welfare. Animal experiments were performed in accordance with the "Guidelines for animal experimentation" of the Aomori University of Health and Welfare.
1. Materials and rearing environment
Animals were male Wistar rats (n=24) with a postnatal age of 28 days, randomly divided into a control (CON) group (n=8), a running exercise (RUN) group (n=8), and a jumping exercise (JUM) group (n=8). All animals were acclimatized for one week, with temperature and humidity at 23±1℃ and 55±5%, respectively, throughout the rearing period; their dark period was 12 hr from 7 pm to 7 am. During rearing, animals were allowed to drink tap water and consume CE2 (CLEA Japan Inc., Tokyo, Japan) feed for laboratory animals, ad libitum, and were also free to move and engage in physical activity in their cages.
2. Training protocol
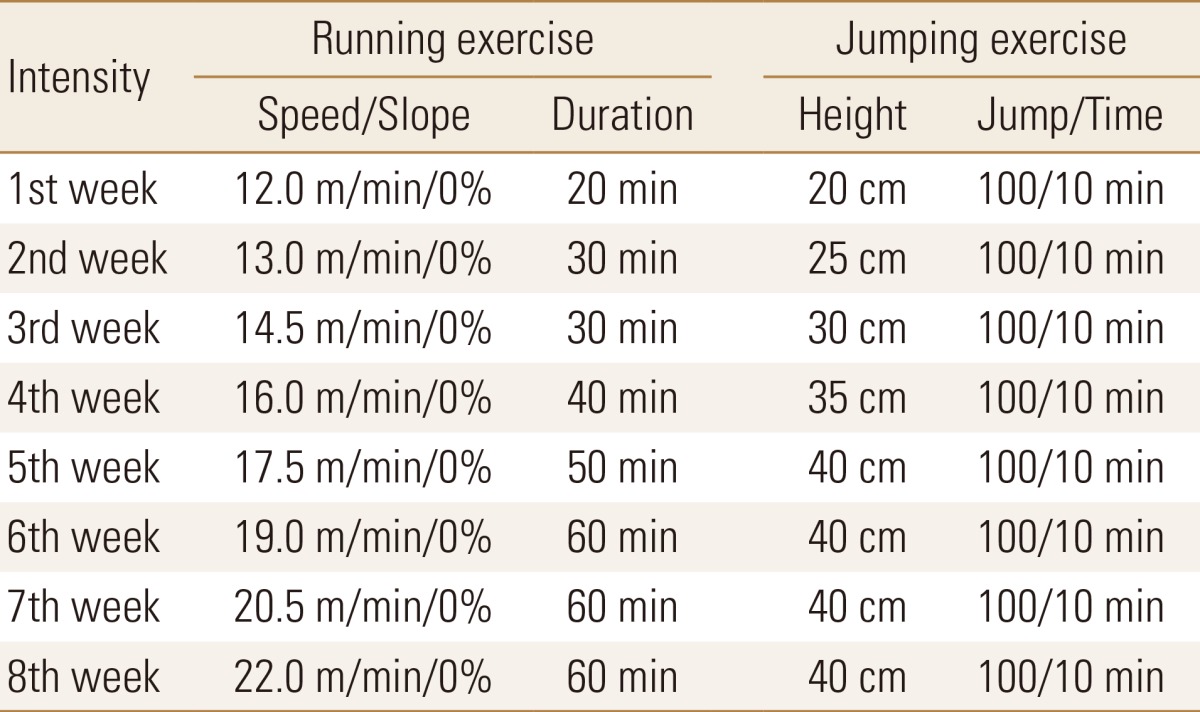
In the RUN and JUM group, three days of preliminary exercise were performed in order to prevent the effects of electrical stimulation that occurs during an exercise load. The experiment was begun after 96 hr of rest following the end of the preliminary training. Exercise was done 5 days/week, and was implemented at the same hr for eight weeks.
3. RUN
RUN for rats & mice MK-680 (Muromachi-Kikai, Tokyo, Japan) was used for the RUN group. For exercise load amount, the protocol for aerobic exercise load level proposed by Huang et al.[20] was used, starting from 20 min of exercise at an incline of 0% and speed of 12.0 m/min. Next, the speed and exercise time were gradually increased to an exercise speed of 22 m/min and an exercise time of 60 min (Table 1).
4. JUM
We employed the preceding method of Umemura et al. [21] and Sueda et al.[22] for the JUM Group, and made the subjects perform a type of anaerobic exercise i.e., jumping. For the JUM, a box surrounded by four boards of 25 cm in diameter and 20 cm in height was created in accordance with the method by, and an electrical stimulation device was installed on the floor. Next, the rats were enabled to jump spontaneously from the floor, with one jump being understood to be when the forefeet hit the upper edge of the four-sided enclosure. The exercise load started from 100 times/10 min and a height of 20 cm, and then increased by 5 cm height increments every week, finally reaching a maximum of 40 cm on the four weeks (Table 1).
5. Blood component
Each group had blood taken from the caudal vein immediately before and immediately after the exercise loading, to measure blood lactic acid levels and bone metabolism markers.
6. Bone metabolism markers
Blood was drawn from the tail caudal vein on the 4th week and the 12th week, at rest and immediately after exercise stress. Taking into consideration the in-day fluctuations of bone metabolism markers for rats, blood-collection was performed in the afternoon (pm 4-6). The collected blood was centrifuged for 15 min at 3,000 rpm and the separated plasma was tested for bone metabolism markers. We measured plasma osteocalcin (OC) level, an osteogenetic factor, using a Rat OC EIA Kit (Biomedical Technologies Inc., Stoughton, MA, USA). Blood levels of tartrate-resistant acid phosphatase (TRACP), a bone absorption factor, were measured using an acid phosphatases colorimetric assay kit (Cayman Chemical Co., Ann Arbor, MI, USA).
7. Bone morphogenic
1) Bone mass and bone length
After the experiment was completed, a thoracotomy was performed under pentobarbital sodium (50 mg/kg) anesthesia, the rats were slaughtered by perfusion with physiological saline, and the femora and tibiae were immediately extracted. Total maximum bone length was measured three times using electronic calipers ABSOLUTE Digimatic CD-S20C (Mitutoyo Corp., Kanagawa, Japan), and the average value was recorded as the bone length. After the muscle tissue was removed, the wet weight of each bone was measured using an electronic balance. The bones that underwent morphological measurements were then subjected to a three-point bending test. They were subsequently soaked for 5 days in a mixed solution of chloroform and methanol and dried for 1 hr at 120℃, at which time their defatted dry weight was measured.[22]
8. Bone strength
The bones subjected to morphological measurement were covered with gauze moistened with physiological saline to prevent drying. A three-point bending test was performed using an Instron Model 4465 tensile tester (Instron Co., Norwood, MA, USA) bone fracture measurement instrument, and results were analyzed using Bluehill II. In the three-point bending test, pressure was applied from above and below to one point in the center of the bone and the maximum bending load was measured. Diaphysis center measurements were performed at the maximum bending load using a span of 16 mm and a rate of crosshead speed of 10 mm/min.[23]
RESULTS
1. Body weight
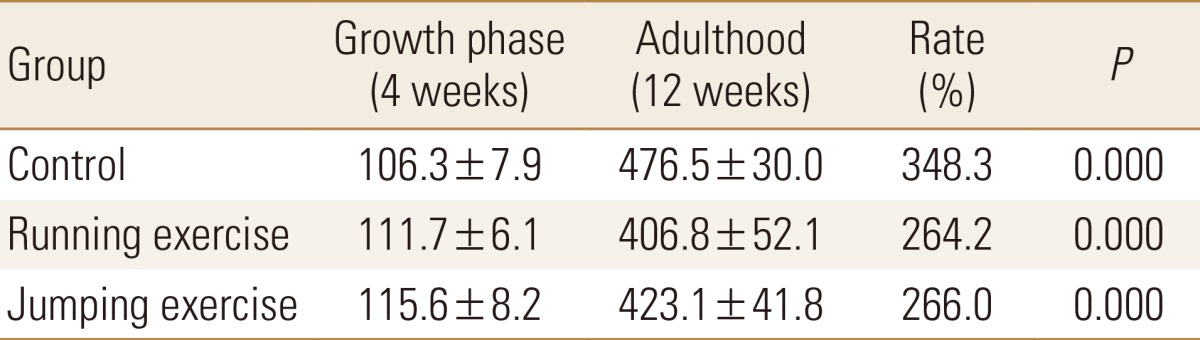
Compared to that at the start of the experiment, weight had increased significantly at the end of the experiment in all the groups, at 348.3% for CON group, 264.2% for RUN group, and 266.0% for JUM group (P<0.001, in each). The mean bone weight in the RUN group was significantly lower than that in the CON group after the experiment was completed (P<0.01, Table 2).
2. Change in bone metabolic factors due to mechanical stress
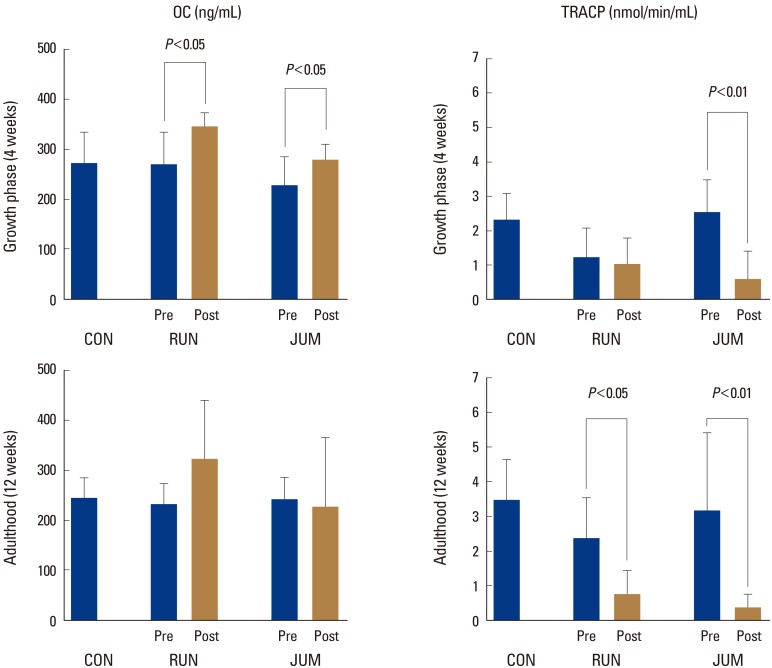
OC in the growth phase increased significantly after exercise loading at a rate of 36.3% in the RUN group and 29.0% in the JUM group (P<0.05, in each) (Fig. 1).
On the other hand, TRACP decreased significantly after exercise loading in the JUM group only at a rate of 77.0% (P<0.01). In contrast, TRACP in adulthood decreased significantly after exercise loading compared to before at a rate of 71.6% (P<0.05) in the RUN group and 86.0% (P<0.01) in the JUM group (Fig. 1).
3. Bone metabolism marks at rest
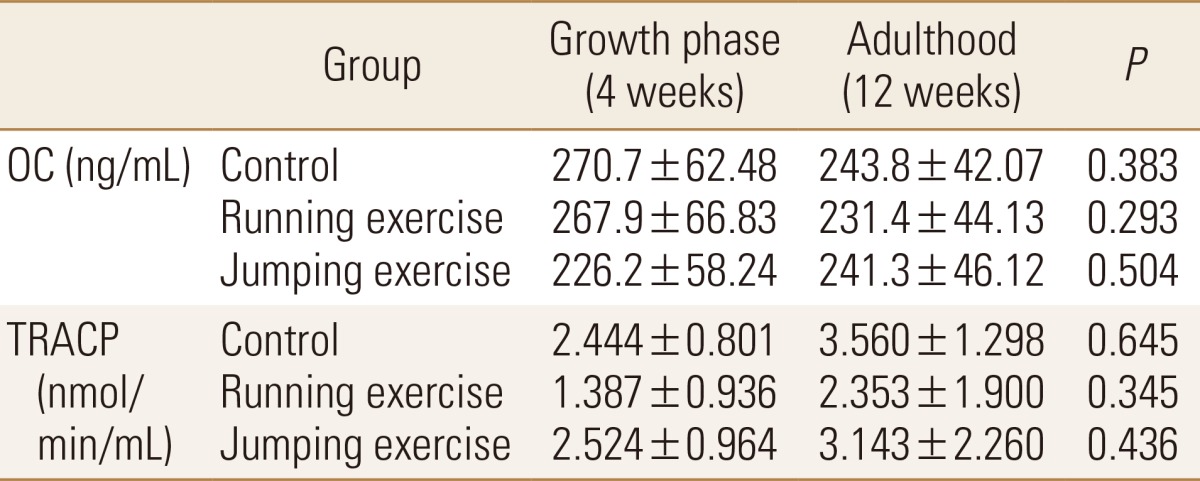
Bone metabolism markers at rest in the growth period and at rest in the adulthood and without significant difference in all groups (Table 3).
4. Morphology of bone
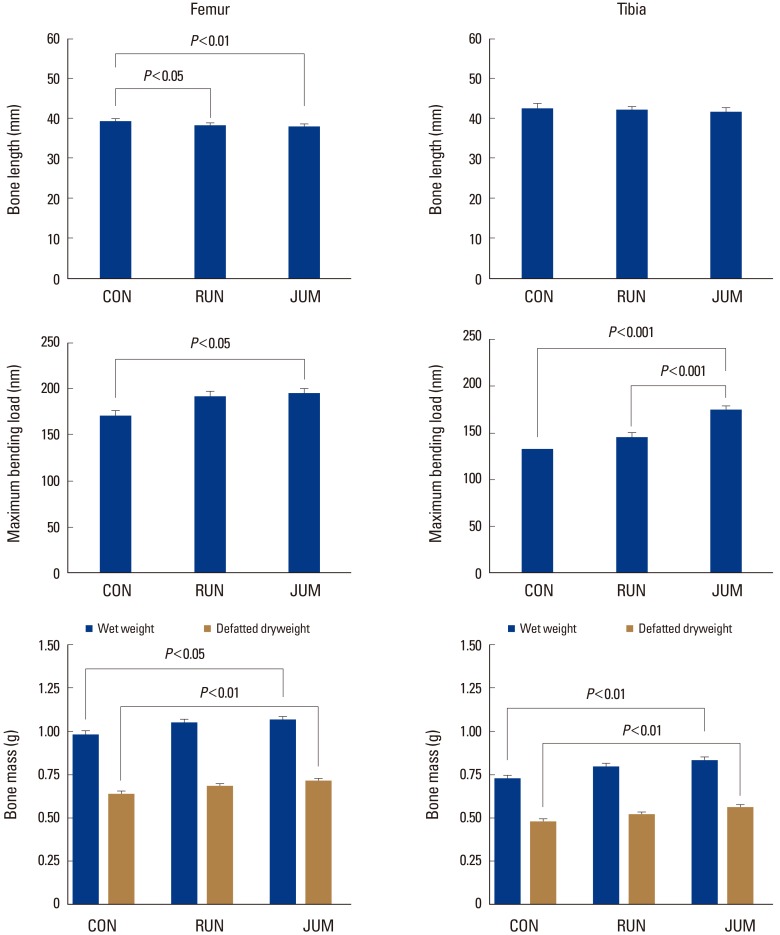
The maximum bone length of the femur was significantly shorter in the RUN (2.1%, P<0.05) and JUM (3.1%, P<0.01) groups compared to the CON group (Fig. 2).
The maximum bending load of the femur was significantly higher in the JUM group (14.5%, P<0.05) than in the CON group, while that of the tibia was significantly higher in the JUM (30.7%, P<0.001) and RUN (19.9%, P<0.001) groups than in the CON group (Fig. 2).
Wet weight and defatted dry weight of the femur were significantly higher in the JUM group than in the CON group at 8.4% (P<0.05) and 11.8% (P<0.01), respectively. Wet and defatted dry weights of the tibia were also significantly higher in the JUM group than in the CON group at 14.9% and 17.6%, respectively (P<0.01, in each) (Fig. 2).
DISCUSSION
In this study, we examined the effect of exercise loading characteristics on bone metabolic response and bone morphology in the growth phase and adulthood. With regard to bone metabolism in response to exercise loading, we found that in the growth phase, modeling was appreciably activated primarily through bone morphogenetic factors, whereas in adulthood, exercise loading contributed to pronounced suppression of bone resorption factors. Additionally, intense exercise loading suppressed bone resorption factors regardless of animal age, while bone metabolism factors responded only to transient exercise loading. On the other hand, bone mass and bone strength in adulthood were proportional to exercise loading, whereas bone length was inversely proportional to exercise loading.
Mechanical stress is a bone formation-promoting factor that is fundamental to the living body as well as an important mechanism of increasing bone mass.[24,25,26] Frost proposed two processes for bone metabolism kinetics accompanying mechanical stimulation, namely modeling and remodeling.[19] Depending on magnitude, mechanical strain applied to bones is classified as disuse window, physiological window, mild overuse window, and damage window. Disuse window induced acceleration of bone resorption has been reported.[19,27] Osteocytes respond to even slight strain due to mechanical stress,[28] and if bone strain exceeds a certain level, osteoclasts are mobilized and the remodeling cycle occurs.[29]
However, bone morphogenetic factors increased in the growth phase regardless of the exercise load quantity, while bone resorption factors decreased only in response to an intense load. On the other hand, in adulthood, bone morphogenetic factors did not change despite exercise loading, and only bone resorption factors were suppressed by exercise loading. Bone modeling or remodeling occurs due to mechanical stress or microdamage. Furthermore, at the stage of significant osteogenesis in the growth phase, bone tissue quantity increases due to preferential modeling activity and is adapted to bone morphology according to mechanical demand.[30] Mechanical stress through exercise load is an effective way to increase one's bone density and bone mass, and a strong exercise load contributes to greater bone improvements.[16,17,31,32] Morey and Baylink [33] reported no change in bone resorption factors in zero gravity and that the adaptation to bone morphology is enhanced according to mechanical demands. If we consider our results with regard to the mechanical levels of Frost, the exercise load quantity of the JUM group is the level at which microdamage occurs. In other words, we believe that bone metabolic response to the load quantity used in the JUM group temporarily suppresses the remodeling that accompanies cell repair.
Increases in the blood concentrations of the bone-related factors dehydroepiandrosterone (DHEA) and estrogen have been reported immediately after medium- and high-intensity exercise loads, and Aizawa et al.[34] implicated that increased DHEA levels affect bone-related factors.[35] In bone metabolism in the growth phase, modeling that enhances bone morphogenetic factors is selectively dominant, and the changes in bone metabolism due to exercise loading seen in this study support the results of previous studies.[30,34,35] Osteocytes that undergo Mechanical stress are regulated by osteoblasts and osteoclasts.[36] Further, bone tissue mass is determined by a balance inside the body based on the magnitude of the effects on osteoblasts and osteoclasts.[37] On the other hand, it has also been reported that no significant change in bone-associated factors was seen beyond 4 to 5 weeks after the start of exercise loading, and the possibility of a change in the threshold value at which a bone responds to Mechanical stress has been suggested.[38] Shimamura et al.[39] demonstrated that bone mass acquired through exercise is lost when exercise is discontinued and suggested that continuous exercise loading is necessary to maintain bone mass. In this study, no significant change in resting bone metabolism was seen between the growth phase and adulthood. That is, we believe that there is little permanent effect of Mechanical stress on bone metabolic factors and that the suppression of bone resorption factors is dependent on transient and intense Mechanical stress.
With regard to bone morphology, Nishioka et al.[40] suggested the suppression of long bone growth along the major axis due to exercise loading from an early age. Additionally, in a previous study in which exercise loading was performed during the growth phase, de Campos-Ferraz et al.[41] also reported suppression of bone growth along the major axis and a short body length. On the other hand, Sueda et al.[22] reported that jumping 50 times per day resulted in no change in femur length but did result in a higher bone mass and circumference.
In this study, maximum bone length exhibited low values in proportion to exercise load quantity, whereas bone weight and maximum bending load exhibited high values. The exercise load quantity used in our study was more than twice that used by Sueda et al.[22], and the change in lactic acid due to exercise loading exhibited different responses in the growth phase and adulthood. In short, we believe that the threshold level of Mechanical stress that affects bones also changes with growth, but we were unable to determine those threshold values in this study. In summary, bone mass and bone strength in the growth phase increase according to exercise load quantity above a certain level, and continuous excessive exercise load may hinder long bone growth along the major axis.
From these facts, we conclude that transient intense exercise loading in the growth phase induces marked enhancement of bone morphogenetic factors and suppression of bone resorption factors and contributes to PBM and bone strength; however, exercise load quantity should be considered from the perspective of long bone growth along the major axis.
CONCLUSIONS
The bone metabolism factors in this study showed a short-lived reaction to exercise stress, and were significantly influenced by the type of exercise stress applied. Although high-intensity exercise stress during the growth period leads to strengthening of the bone, it carries the risk of suppressing bone length. During rapid bone growth periods, therefore, it is necessary to take into consideration the tradeoff between increased bone strength and bone length, and to apply appropriate exercise stress.
 PDF Links
PDF Links PubReader
PubReader ePub Link
ePub Link Full text via DOI
Full text via DOI Download Citation
Download Citation Print
Print